Les Lépidoptères
rhopalocères et hespéridés
du Bois des Fagnes
(prov. Namur)
(Lepidoptera Rhopalocera & Hesperiidae)
Le Bois des Fagnes
François Delacre,
Ingénieur des Eaux & Forêts
Jean Delacre
« Il ne faut jamais perdre de vue qu’une forêt n’est pas seulement une collection d’arbres économiquement exploitable, c’est avant tout un milieu biologique, l’un des plus complexe de la nature, dont tous les éléments (arbres, arbustes, herbes, sol, micro-organismes, faune) sont interdépendants. On ne peut détruire l’un de ces éléments sans risquer de bouleverser l’équilibre biologique de l’ensemble et de provoquer à plus ou moins brève échéance la disparition de la forêt elle-même »
H. PABOT (1957)
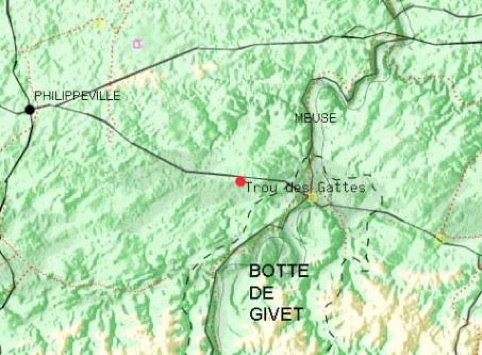
Situation géographique
Le bois des Fagnes fait partie d’une longue bande forestière de la Fagne schisteuse (province de Namur - Belgique), qui s’étire depuis Agimont à l’est jusqu’à Roly et Fagnolle à l’ouest. Il s’étend sur les communes de Romedenne, Vodelée, Gimnée, Doische et Agimont.
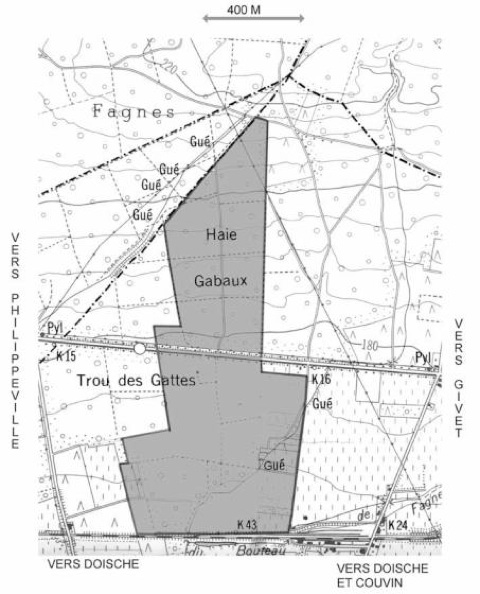
Le territoire étudié dans le cadre de cette étude (recensement des Rhopalocères) est plus restreint et ne concerne qu’une propriété privée (Trou des Gattes) de 40 hectares sur la commune de Doische et la Réserve Naturelle RNOB de la Haie Gabaux de 30 hectares.
Topographie
Le bois des Fagnes est situé dans la dépression famenienne et son relief est très doux, légèrement incliné vers le sud ( pente de moins de 5°).
Son altitude varie de 170 m au sud à 225 m au nord. Des prairies (grasses, maigres ou marécageuses) se situent de part et d’autre de la langue de forêt.
Deux ruisseaux à débit temporaire confluent au sud du territoire tandis qu’un troisième, également intermittent, prend sa source dans le nord de celui-ci avant de le quitter. Tous trois font partie du bassin hydrographique du ruisseau de la Joncquière.
Climat
La température moyenne annuelle est de 8,6°C, et les précipitations moyennes s’élèvent à 781 l/m2 l’an (VAN DIEPENBEEK, in litt.). Celles-ci semblent bien réparties, mais les argiles schisteuses sont pauvres en eau utile et ont une mauvaise conductivité hydrodynamique. Elles s’assèchent très vite et peuvent même se crevasser durant les étés secs (NOIRFALISE, 1984).
Il gèle en moyenne pendant 94 jours (donnée des années 80/90 !). Les dates moyennes de la première et de la dernière neige sont le 17 novembre et le 10 avril (VAN DIEPENBEEK, in litt.).
La période de végétation s’étend sur un peu moins de six mois. Le climat est tempéré humide à hiver relativement doux et à été frais.
Géologie
D’après FORIR (1989), le groupe Primaire est représenté par le Dévonien supérieur.
L’Étage famennien (Fa) est représenté par l’Assise de Mariembourg : schistes souvent violacés avec psammites (Falb) et surtout par l’Assise de Senzeille : schistes souvent verdâtres, fréquemment noduleux (Fala).
L’Étage frasnien occupe la pointe nord du territoire avec des schistes divers, assez souvent noduleux (Frlm).
Les schistes famenniens ont donné beaucoup de sols lourds et peu épais (AVRIL, 1990).
Pédologie
Dans les zones planes, notamment sur le Famennien, le schiste s’est altéré en argile lourde, panachée, donnant des sols à drainage imparfait. Il se forme par temps humide une nappe perchée qui percole lentement dans le niveau plus lourd (substrat d’argile d’altération) qui apparaît à moins d’un mètre. Cette nappe perchée et plus ou moins temporaire, à cause d’un drainage naturel défectueux, explique l’utilisation presque exclusive de ces sols plats par la prairie ou la forêt (AVRIL, 1990).
Les différents types de sols rencontrés au Bois des Fagnes sont des sols limoneux fortement gleyifiés (*) à développement de profil non défini et substrat schisteux débutant à faible profondeur (fAxh), des sols fortement gleyifiés sur matériaux limoneux (Ahp), des sols argileux fortement gleyifiés à développement de profil non défini (Ehx), des sols argileux modérément gleyifiés à horizon B structural (Edb), des sols limoneux à horizon B structural et charge schisteuse (Gbbf) ou peu caillouteux (Gbb), et des sols limoneux peu caillouteux modéréments gléyifiés à horizon B structural (Gdb) (AVRIL, 1984 et 1990).
Le type de sol dominant est Edb, sol brun acide, dont voici la description (AVRIL 1984) :
De 0 à 0,2 cm : feuilles de chênes et herbes partiellement décomposées ; brun très foncé.
De 0,2 à 8/12 cm : argile limoneuse brun grisâtre très foncé ; structure grumeleuse et subpolyédrique prononcée ; légèrement dure ; beaucoup de petites traces de rouille ; limite inférieure distincte et ondulée.
De 8/12 à 34 cm : argile limoneuse brun pâle ; structure subpolyédrique (**) et polyédrique prononcée dure ; quelques graviers schisteux ; coatings d’argile peu distincts et discontinus ; gleyification peu distincte ; quelques traces de rouille et inclusions de Fe et Mn ; limite inférieure distincte.
De 34 à 62 cm : argile lourde grise à gris clair ; structure polyédrique (***) prononcée ; extrêmement dure ; argile d’altération de schistes renfermant peu de graviers schisteux ; gleyification distincte ; la teinte du fond est grisâtre avec beaucoup de petites taches de rouille distinctes ; limite inférieure abrupte.
De 62 cm et + : schistes famenniens non altérés.
(*) Gléyfication = phénomène de réduction du fer, provoqué par l'absence d'oxygène, dans la zone de sol à nappe phréatique permanente (en toute saisons). Lorsque l'excès d'eau dans le sol (l'hydromorphie) est temporaire, il se traduit par une oxydation du fer et donc la présence de taches de rouille. Le gley est la zone gris-bleu et le pseudo-gley est la zone bariolée avec alternance de taches grises et de taches de rouille; le pseudogley correspond à la zone de battement de la nappe (régime hydrique alternatif).
(**) Structure subpolyédrique = structure presque en polyèdre
(***) Structure polyédrique = structure des argiles en polyèdre (solide limité de toutes parts par des polygones plans)
Phytosociologie
Le Bois des Fagnes se range dans la classe du Querco-Fagetea, dans l’ordre du Fagetalia silvaticae, dans l’alliance du Carpinion betuli (chênaies
mixtes à Charmes) et dans l’association du Stellario-Carpinetum (chênaie-charmaie à Stellaire) (NOIRFALISE, 1984).
Le Stellario-Carpinetum est caractérisé par le groupe du Charme et de la Stellaire holostée et par le groupe de l’Anémone sylvie. C’est une association acidocline, sur sols bruns lessivés et bruns acides. Le peuplement principal est formé par le Chêne pédonculé et le Chêne rouvre. Le sous-bois ou les taillis comportent principalement le Charme, le Coudrier, les chênes, l’Érable sycomore, le Hêtre et des espèces silicicoles (Bouleau, Tremble, Sorbier des oiseleurs, Bourdaine).
La végétation herbacée est marquée par l’apparition de transgressives du Quercion (groupe de la Houlque et groupe Muguet) (NOIRFALISE , 1984)
La sous-association du Bois des Fagnes est le stellario-Carpinetum-caricetum (chênaie-charmaie schisteuse), caractérisée par le groupe du Carex flacca. Les épineux y sont représentés par le Prunellier et les Aubépines (NOIRFALISE, 1984).
Régime forestier
Le Bois des Fagnes est un taillis sous futaie. Le Chêne pédonculé [Quercus robur L.] est largement dominant. Il est accompagné du Chêne sessile [Quercus petraea MATTUS], du Bouleau verruqueux [Betula pendula ROTH], du Bouleau pubescent [Betula pubescens EHRN.], du Charme [Carpinus betulus L.], du Peuplier tremble [Populus tremula L.] et de quelques Frênes [Fraximus excelsior L.]. Quelques enclaves sont constituées d’une futaie claire de Chênes pédonculés et de Chênes rouvres.
Le taillis pur n’occupe que quelques hectares et des plantations résiduelles d’épiceas couvrent actuellement trois hectares et demi et ne seront pas remplacées. La régénération artificielle est désormais axée vers les feuillus : Chênes, Alisiers, Merisiers, Érables, Frênes… Les résineux (Douglas, Pin sylvestre, Mélèzes) sont introduits en îlots dans certains secteurs très pauvres.
À l’extrême sud de la propriété, dans une étroite bande de terrain hors Famennien, une aulnaie naturelle [Alnus glutinosa (L.) GAERTN.] a été récemment éclaircie pour mettre en valeur les meilleurs fûts.
Disséminés dans toute la forêt, se trouvent quelques Pommiers sauvages [Malus sylvestris MILL.], quelques Alisiers [Sorbus torminale (L.) CRANTZ], quelques Sorbiers des oiseleurs [Sorbus aucuparia L.], et quelques Viornes obier [Viburnum opulus L.].
Le Hêtre est absent à quelques rares pieds près d’ailleurs rabougris, tandis que le long de la route principale (Philippeville-Givet), quelques Érables, Frênes et Ormes semblent provenir de semis naturels provenant des arbres d’alignement.
Les lisières de la forêt sont colonisées par le Prunellier [Prunus spinosa L.], l’Aubépine à un style [Crataegus monogyna JACQ.], l’Aubépine épineuse [Crataegus laevigata (POIR.)DC.] et différents Saules [Salix sp.].
Impact des aménagements forestiers sur les Rhopalocères
de la Haie Gabaux / Trou des Gattes
(Bois des Fagnes - Doische)
Jean Delacre
Vocations pionnières de la propriété
Jusqu’au milieu du XIXème siècle, l’ensemble de l’aire étudiée était constitué de prairies marécageuses drainées par un réseau de fossés creusés de mains d’homme (dont certains témoins apparaissent encore çà et là dans l’actuelle forêt) ou de landes à bruyère à l’usage de pâturage collectif pour les troupeaux du village.
Avec l’abandon progressif de ce pacage, sans nul doute en raison de sa piètre qualité, ces prairies et ces landes furent lentement recolonisées par la forêt qui y reprit ses droits. Aucun chêne n’y dépassant l’âge de 120/125 ans, cela nous documente approximativement sur l’époque de la reconquête forestière.
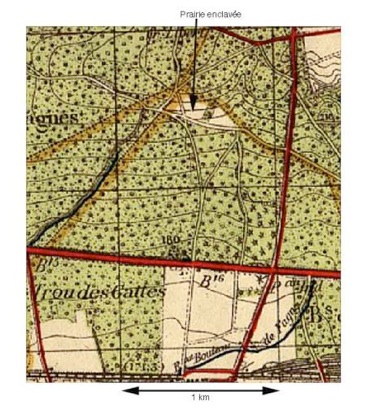
Une carte de l’Institut Géographique National datant de 1941 nous démontre l’existence à cette époque d’une dernière prairie enclavée de 8 à 9 hectares, au nord de la propriété déjà totalement recolonisée par les épineux vers 1955 d’après mes souvenirs d’enfance, puis par la forêt mixte composée de Prunelliers , Aubépines, Chênes et Bouleaux telle qu’elle se présente aujourd'hui. Depuis les années 1880, à l’exception de cette prairie recolonisée tout récemment, la gestion n’a donc plus été l’objet des éleveurs et agriculteurs mais celle des forestiers et des chasseurs.
Tentative d’historique des aménagements forestiers depuis 1923
Entre 1923, année de l’achat de la propriété par mon grand-père Gustave, et 1990, la gestion forestière était avant tout axée sur la chasse. Les essences à croissance rapide (résineux et peupliers) y sont restées de présence discrète. Les « nailles »* étaient tracées pour l’organisation des battues et les coupes de taillis se faisaient plus pour créer des gagnages pour le gibier que pour en retirer un profit. À partir de 1990, la gestion par mon fils François, ingénieur des Eaux & Forêts, sera plus subtile, tant pour mettre en valeur les arbres les mieux représentés (chênes), que pour redonner à la forêt toute sa diversité végétale. Un louable déploiement d’efforts à l’égard d’arbres plus rares, présents sur le site mais jusqu’ici dédaignés lors des coupes de taillis, tels qu’entre autres les Alisiers et les Aulnes glutineux ou les Frênes, a permis d’optimiser la biodiversité et de redonner ainsi à la forêt tout son attrait naturel.
La pratique de coupes de taillis a été poursuivie en rotations de petites parcelles de 30 ares à 2 à 3 hectares l’an, tant pour perpétrer la politique de création de gagnages pour le Chevreuil, que pour créer des parts de bois de chauffage, éclaircir la futaie et favoriser le développement des chênes de qualité. À partir de l’hiver 2001/2002, délaissant l’aspect cynégétique, nous avons innové des aménagements susceptibles de favoriser la survie d’une colonie d’Euphydryas aurinia (le Damier de la Succise), papillon Nymphalinae en grande difficulté en Wallonie et dont les espaces ouverts de cette forêt révèlent constituer un habitat électif . Ce programme sera poursuivi dans les années à venir de façon à obtenir un étagement dans la hauteur des rejets de taillis qu’il faudra ainsi recéper tous les 4 à 5 ans. Il s’agit là d’un réel changement de cap dans les préoccupations des gestionnaires de l’espace.
-
*« naille » en terme de chasse = layon = chemin forestier
Impact de la gestion cynégétique sur les populations de Rhopalocères
Si seules huit à dix populations d’Euphydryas aurinia semblent subsister en Wallonie, pourquoi ce papillon (et bien d’autres) s’est-il maintenu en abondance dans cette partie du bois des Fagnes ? La présence de sa plante-hôte ne serait qu’un des éléments de réponse, car ce Damier a disparu de bien d’autres localisations où pousse encore la Succise.
Quelles seraient les raisons qui ont alors favorisé le maintien d’E.aurinia à la Haie Gabaux / Trou des Gattes jusqu’à nos jours ? Un paramètre favorable est sans nul doute l’apport de lumière (et donc de chaleur) maintenu grâce aux coupes de taillis en rotations annuelles, en une multitude de petites clairières et trouées dispersées dans toute la propriété, ainsi qu’à l’entretien des chemins et des layons structurant l’espace. L’effet de lisière ainsi amélioré semble extrêmement bénéfique pour la petite faune, toujours mieux représentée en orée.
Un autre facteur notable pour le maintien de la meilleure diversité biologique possible est la carence absolue en substances agrochimiques dont a bénéficié tout le site, sa flore et sa faunule.
La trilogie « plante nourricière, lumière et habitat préservé » semble être la bonne recette non seulement pour garantir le maintien, mais aussi pour valoriser l’épanouissement des populations de Rhopalocères forestiers.
Nullement obnubilés par le strict rendement forestier, c’est en favorisant la gestion cynégétique que nous avons, peut-être en toute naïveté, fait du domaine de la Haie Gabaux un havre entomologique et un pays de cocagne pour les papillons, puisque 61 espèces y furent recensées en quatre saisons sur une surface somme toute assez restreinte. Et trois à quatre espèces potentielles comme, Argynnis aglaja, Limenitis populi, Plebeius argus, Cupido minimus, et Satyrium w-album, … y sont encore pressenties.
Chaque cas étant particulier, il faut malheureusement admettre que l’impact « cynégétique » peut être parfois plus désastreux que bénéfique sur les populations de Rhopalocères. Nous venons d’observer à quelques kilomètres de chez nous, ce qu’une prairie « à Succise », retournée et réensemencée par des topinembours (nourriture sur pied pour maintenir – ou attirer ? - les sangliers en forêt) a pu amener comme dégâts sur la population d’E. aurinia connue comme la plus florissante de Wallonie, il y a dix ans à peine !
Les gagnages « engraissés », les semis de plantes fourragères « allochtones » en monoculture, les champs de maïs « traités », ont un impact très négatif sur la faunule. Il n’y a donc pas que du positif dans ces gestions dont la chasse est le lobby.
L’activité agricole a évidemment un impact encore plus néfaste sur les Lépidoptères, à fortiori dans nos contrées où agricole est synonyme d’intensif avec quotas ! Nous avons en bordure de propriété 6 hectares de prairies louées en bail à ferme. L’herbe y est certes belle (enfin dans l’esprit des agriculteurs…), mais quel désert agraire, quel univers abiotique ! L’apport d’engrais, de lisier et d’herbicide sélectif, ainsi que la fauche précoce et répétitive sont de vrais fléaux et sonnent toujours le glas pour la flore, la faune et le futur.
La pratique acharnée de la fauche, en forêt comme d’ailleurs en prairie, est désastreuse pour les Lépidoptères, comme pour les Reptiles, les Batraciens et les autres occupants de ces lieux ouverts. L’exercice de la chasse exige des chemins et des bermes bien dégagés avant tout pour une question de visibilité et de sécurité. Ces layons sont donc gyrobroyés chaque année après la floraison estivale, et suffisamment tard pour qu’il n’y ait qu’un regain limité au moment des battues d’octobre / novembre. L’habitude des gestionnaires de chasse est de passer la « tondeuse » en juillet ou en août. C’est assurément très destructeur pour le cycle des papillons dont les chenilles ne sont pas encore à cette époque, ni entrées en diapause, ni chrysalidées, ou encore néonates, voire les œufs candidats à l’hibernation très exposés.
Pour ne pas qu’un massacre au tracteur ou autre machine agressante ne détruise d’une part des milliers d’années de genèse, d’autre part les efforts (très récents…) de quelques propriétaires et acteurs convaincus du bien fondé des directives de Natura 2000, ici pour Euphydryas aurinia, ailleurs pour d’autres espèces fragiles, il semble que l’essentiel de ce qui a été dit (voire dicté…) reste à faire. Un cahier des charges aux règles pointilleuses et à la surveillance accrue devrait faire l’objet d’une meilleure concertation entre les propriétaires forestiers (état, communes, particuliers) et les usagers (agriculteurs, forestiers, chasseurs), et ce, dans le cadre des contrats de gestion Natura 2000.
Il n’est pas outrancier d’imaginer plausible le moyen de concilier la préservation de l’entomofaune, de l’herpétofaune et des autres locataires prioritaires de l’humus des layons forestiers, avec le nécessaire besoin de sécurité exigé par les chasseurs, qui, ne l’oublions pas apportent aux propriétaires forestiers (tant privés que communaux ou domaniaux), un apport de devises non négligeable. Les fauchaisons devraient être mises en œuvre en octobre à vingt centimètres du sol pour respecter les plantes, les chenilles et la petite faune et faunule. Gyrobroyer brutalement les chemins forestiers jusqu’à la racine des plantes, et des herbages est désormais inconciliable avec la préservation de la biodiversité et les exigences écoconscientes induites par la légitime politique du développement durable. Si l’entomologiste accepte la nouvelle éthique du moindre prélèvement, il reste pantois devant l’anéantissement du biotope froidement perpétré sous ses yeux par des pratiques imbéciles mais souvent légalisées. D’autant plus que le premier cité n’est qu’un bien petit et respectable prédateur alors que le second se rend le plus souvent coupable d’un triste et irréversible saccage.
Natura 2000 et les aménagements futurs
Sensibilisé au devenir des sites d’intérêt biologique, et des Lépidoptères en particulier, contrairement à bon nombre de propriétaires forestiers plutôt réservés vis-à-vis de Natura 2000, j’ai œuvré pour que le Bois des Fagnes soit sélectionné.
Conseillé par le biologiste Philippe GOFFART (chercheur à l’Observatoire de la Faune, de la Flore et des Habitats (OFFH ) au Centre de Recherche de la Nature, des Forêts et Bois, à Gembloux), j’ai anticipé sur les travaux qui me semblaient souhaitables dans le cadre des contrats de Gestion Natura 2000. Pour préserver dans l’urgence un habitat acceptable pour Euphydryas aurinia et son cortège, j’ai amorcé des travaux d’aménagement ciblés sur cette espèce.
Les « layons à Succise » seront progressivement élargis à raison de 100 à 300 mètres l’an, en un maillage continu (élargissement sur 15 à 20 mètres, aussi bien dans l’axe nord/sud que est/ouest) tout en laissant sur pied quelques plants et arbustes, comme l’Aubépine, le Pommier sauvage, le Prunellier,et quelques chênes isolés, tout en veillant à ce que la lumière puisse partout inonder ces chemins et y apporter la chaleur nécessaire à l’épanouissement des plantes florifères (et des chenilles). Un recépage devrait être élaboré tous les 4 à 6 ans, pour empêcher la forêt de regagner du terrain, tout en respectant certaines essences basses . Ce maillage devrait être ouvert sur les plaines, et sur le parcours de la ligne de haute tension qui traverse la propriété ; celle-ci aurait dù être recépée tous les quatre à cinq ans, mais s’est malheureusement totalement refermée après 14/15 ans faute de travaux de dégagements . Les responsables Natura 2000 devront sensibiliser la société ELIA*, pour que les plans initiaux soient respectés. Dans les premières années suivant la mise à blanc, c’était devenu une véritable autoroute à papillons, une aire de vol d’autant plus utile, qu’elle favorisait la dispersion des « divagants » qui pouvaient ainsi recoloniser d’autres sites, tout en gardant de petits réservoirs génétiques prêts à pallier à des disparitions localisées.
(* Chose faite : en 2006, nous avons signé un contrat de gestion « écologique » avec Elia, pour que la ligne de haute tension qui a été réouverte sur une largeur de 50 mètres, soit fauchée un an sur deux pour favoriser la présence d’Euphydryas aurinia autant d’ailleurs que celle des reptiles présents sur le site)
Grâce aux travaux que nous avons commencés l’hiver 2001/2002, nous avons déjà pu revoir au printemps 2003 Iphiclides podalirius butinant les fleurs d’Aubépines laissées en place. C’est de bon augure pour l’avenir, car il n’avait plus été aperçu sur le site après 1980.
Au printemps 2003 et plus encore en 2004, notre cher Damier de la Succise volait sur tous les chemins aménagés et dans toutes les clairières à Succise : franc encouragement pour persister dans cette voie !
La propriété de la Haie Gabaux - Trou des Gattes - Crestia, est reprise dans l'inventaire des Sites de Grand Intérêt Biologique (SGIB) sur le site internet de la DGRNE (Informations sur le Biodiversité en Wallonie - Observatoire de la Faune, de la Flore et des Habitats).
En juin 2004, la Fondation Wallonne pour la Préservation des Habitats, portait à notre connaissance par une lettre de son président le Baron Edgar Kesteloot que le Prix Interbrew pour l’Environnement 2004 nous était décerné pour les efforts que nous avions déployés pour la survie d’Euphydryas aurinia dans notre propriété, prix qui nous a été remis par S.A.R.le Prince Laurent de Belgique le 21 septembre 2004.
Observations commentées
sur la faune des LÉPIDOPTÈRES rhopalocères & hespéridés
du Trou des Gattes / Haie Gabaux à Doische.
(Fagnes – Province de Namur - Belgique)
Jean DELACRE (2001/2004)
INTRODUCTION
La présente étude est le fruit d’une centaine de sorties étalées sur quatre années (2001/2004) de janvier à septembre. Les macro-photographies ont été réalisées avec un boîtier Minolta Dynax 5, un objectif macro 100 mm à ouverture 2.8 et un flash spécial macro à bras télescopiques de marque Minolta (Macro twin flash 2400). Les photos ont été scannées sur un Nikon Coolscan IV ED. Les films utilisés sont en majorité des diapositives Fuji Sensia 100 et 200 ASA, et parfois Fuji Provia 400 ASA pour les prises de vue hivernales.
Pour des raisons de cohérence et de facilité, je me suis volontairement limité à une aire très restreinte (+/- 70 hectares). J’ai tenté de réaliser au mieux le recensement de la faune des Rhopalocères évoluant sur ce territoire privé du moins pendant la période de cette étude (30 hectares en R.N.RNOB à partir de 2009), tout en essayant de mener une étude de l’impact de la gestion forestière et cynégétique sur les Lépidoptères. Subséquemment à cette étude sur les Rhopalocères du Bois des Fagnes et de concert avec mon épouse Monique, j’ai entamé un pré-inventaire photographique de la flore présente sur le site (MONIQUE DELACRE det.). La situation, la topographie, le climat, la géologie, la pédologie, la phytosociologie et le régime forestier sont co-traités dans le chapitre «Le Bois des Fagnes » avec mon fils François, Ingénieur des Eaux & Forêts.
Toutes les cartes de répartition Wallonnes des Rhopalocères présents sur le site étudié, et placées en tête des photographies de chaque espèce, ont été adaptées et modifiées au départ des travaux de Philippe GOFFART & Benoit DE BAST (2000). Les cartes originales sont consultables sur le site de la DGRNE .
Ces cartes, qui seront mises à jour en fonction de l'état d'avancement des informations récoltées par les observateurs de terrain du Groupe de Travail Lépidoptères (GTL), dont je suis un modeste collaborateur “amateur” sont compilées, encodées et archivées au Centre de Recherche de la Nature, des Forêts et du Bois à Gembloux. Aucune carte de ce genre ne peut évidemment être considérée comme définitive et des lacunes seront malheureusement toujours présentes et difficilement comblées par manque d’informations ou de prospection dans des zones de "moindre intérêt" ( du moins pour les lépidoptéristes...). Leur but est de donner une approche (très relative) de la présence de telle ou telle espèce dans une aire donnée, de mettre en évidence les régions les plus riches en nombre d’espèces, mais certainement pas de signaler une absence, ce qui serait très difficile, voire impossible à prouver.
FAUNE DES RHOPALOCÈRES
Si 114 espèces de Rhopalocères et Hespéridés ont été répertoriés avec certitude en Wallonie (103 autochtones), seulement 92 l’auraient été après 1990 et la région de Fagne/Famenne/Calestienne, l’une des plus riche de Wallonie, représenterait 72 espèces, dont 68 dans la seule maille UTM (Universal Transverse Mercator) de 10 x 10 km concernant ce site et enveloppant toutes les unités stationnelles. (31UFR25)
L’aire étudiée ne représente qu’un seul type d’habitat : celui forestier, incluant lisières, clairières, prairies sylvatiques et allées, sur sol brun acide. Quelques chemins empierrés de longue date peuvent cependant être considérés comme de nature calcaire (une flore spécialisée en témoigne...) et illustrent un second habitat subsidiaire à prendre en considération pour expliquer la présence locale d’espèces calcicoles.
Avoir réussi à répertorier 61 espèces de Rhopalocères sur cet habitat réduit est somme toute un bilan tant inespéré qu’enthousiasmant. Je tente dans le chapitre aménagements d’expliquer ce résultat, par le type d’une gestion forestière particulière dont la continuité fût maintenue depuis des décennies.
HESPERIIDAE
Sept espèces d’Hesperiidae fréquentent le site, mais seules Erynnis tages, Ochlodes venatus, Pyrgus malvae, Thymelicus lineola et Thymelicus sylvestris se rencontrent en abondance.
Carcharodus alceae n’a été aperçu à ce jour qu’une seule fois en 2002 et deux fois en 2003.
Carterocephalus palaemon est assez localisé sur une petite clairière au sein de la forêt, mais est abondant sur ces quelques mètres carrés (première observation, le 11 mai).
PAPILIONIDAE
Deux espèces de « Porte-queues » sont présentes sur le site.
Papilio machaon ne se rencontre qu’en exemplaires isolés et une femelle a été vue en oviposition sur le site à plusieurs reprises en 2002 et 2003. Sa présence était plus ré gulière en 2004.(jusqu’à sept ou huit imagos sur une même prairie)
Quant à Iphiclides podalirius, si j’ai le souvenir de ses nombreuses apparitions dans les années 1950 à 1970, je ne l’avais plus observé après 1980. La découverte en mai 2003, d’un bel imago butinant des fleurs de Crataegus monogyna dans une de nos dernières éclaircies entamées l’hiver 2001/2002 en forêt a démontré que nos efforts de gestion pour la récupération des Lépidoptères n’ont pas été vains.
PIERIDAE
Neuf espèces ont été répertoriées à ce jour.
Gonepteryx rhamni est chaque année l’espèce pionnière, avec par exemple une première apparition en 2003 le 23 février. Le « Citron » est l’espèce prééminente sur le site, avec souvent plusieurs centaines d’individus volant de concert ou butinant dans les clairières florifères. Il faut dire que la Bourdaine, sa plante-hôte est bien représentée dans toute la forêt. Seuls parmi les Pieridae, Colias crocea et Colias alfacariensis ne sont généralement qu’occasionnels. Colias crocea était moins rare en 2003, et même abondant en septembre, mais n’oublions pas que ce fut une année exceptionnelle pour les papillons en Wallonie. Le temps ensoleillé de février à septembre induisit des conditions favorables pour les pontes, avec des éclosions massives d’espèces habituellement peu repérables, ainsi qu’une génération potentielle effective pour les multivoltins. Ce fut le cas pour C.crocea et alfacariensis.
Colias alfacariensis n’a été repéré pour la première fois que le 8 septembre 2003 et confirmé à quatre reprises le 21 septembre, mais ne sera probablement jamais que très occasionnel. Par contre, il est assez abondant sur les pelouses calcaires du secteur de Givet et de Foische (quelques kilomètres à vol de papillon) d’où il a tendance à vagabonder, tout comme Lysandra coridon et Erebia medusa.
Aporia crataegi est toujours très présent, et de nombreuses pontes furent observées sur Crataegus monogyna.
A. cardamines, L. sinapis, P. brassicae, P. napi et P. rapae sont fréquents sur l’ensemble du site. A. cardamines ne s’observe en général qu’aux abords des plaines, mais s’introduit en pleine forêt où le mâle, éminent patrouilleur, se livre à une recherche incessante de la femelle.
NYMPHALIDAE Nymphalinae
Un minimum de vingt et une espèces de Nymphalidae Nymphalinae, à ce jour repérées, hantent le site.
Deux parmi celles-ci méritent qu’on s’y arrête.
Nymphalis antiopa (Le Morio), bien que très rare fut encore observé en juillet 2002.
Je pense même que ce fut l’unique notation de l’année pour toute la Wallonie. Estimé comme en « grand danger d’extinction », sa dernière présence est notée des environs de la Botte de Givet. Moins rare avant 1980, il se manifestait alors chaque année à Doische avec précarité (exemplaires isolés). Il fut très abondant dans la proche région de Revin (Ardennes françaises dans les années 1970.
L’avant-dernière observation remonte à 5 ou 6 ans aux alentours du mois de juillet/août. Fraîchement éclos, se séchant au soleil, posé sur un tronc de chêne à deux mètres du sol, il m’a laissé tout loisir de l’examiner, mais j’étais alors démuni de mon appareil photo. Une course effrénée vers la maison, avec retour au plus vite s’est révélée vaine : plus de Morio ! C’est la seule espèce répertoriée dans cet inventaire qui ne soit pas encore illustrée sur mon site web. L’espoir persiste néanmoins d’y arriver un jour...
Nymphalis polychloros s’est manifesté à trois reprises le 30 mars 2003. Donné pour « vulnérable » dans l’Atlas préliminaire des papillons de jour de Wallonie (publication de décembre 2000) par le « Groupe de travail lépidoptères » (Philippe GOFFART & Benoit DE BAAST), la Grande Tortue ne se rencontre plus qu’avec paucité en Wallonie. Le printemps 2004 s’est montré plus généreux, avec des observations plus nombreuses, allant jusqu’à neuf observations dans la seule journée du 28 mars.
Aglais urticae était très discrète en 2001. Plus fréquente en septembre 2002, la Petite Tortue est devenue quasiment pléthorique en juillet 2003 ou j’ai assisté à un véritable pic démographique.
Apatura iris et Apatura ilia (principalement la forme clytie) attestent d’un bon dynamisme populationnel sur le site, et vers le 20/25 juin de chaque année, il est toujours possible d’en observer quelques dizaines d’exemplaires volant de concert dans certaines clairières, et… sur la terrasse de la maison paternelle située en plein cœur de la forêt, ce qui atteste bien d’une tendance rudérale de ces sympathiques papillons... Chaque jour en juin/juillet, plusieurs sujets vont jusqu’à s’introduire dans la maison pour voleter contre les portes fenêtres vitrées et depuis ma plus tendre enfance ce charmant spectacle m’a toujours ravi.
Des grandes Argynnes, seul Argynnis paphia se rencontre partout en nombre. A. adippe est plus localisé dans les clairières intérieures et ne s’est montré de manière plus soutenue qu’en 2003 et suivantes et A. aglaia est moins fréquent mais présent chaque année sur le site Haie Gabaux.
Brenthis ino, Clossiana selene volent dans tous les chemins quadrillant la forêt et ne sont pas bien difficile à repérer.
Issoria lathonia par contre n’a été observé qu’à une seule reprise en 2002, dans une prairie jouxtant le bois, butinant sur une fleur de Succise, deux fois en 2004.
Si Melitaea diamina n’est pas rare, Mellicta athalia n’aurait montré qu’une timide présence en 2003, mais peut-être ai-je été peu attentif les saisons précédentes et aussi peu apte à les différencier en vol et mon absence en juin 2002 est sans doute la raison de cette carence ... (très abondant depuis 2004 suite aux aménagements)
Euphydryas aurinia est à l’origine d’une véritable histoire passionnelle.
Sa première découverte sur notre site remonte à juin 2000. À cette époque, ce n’était pour moi qu’un papillon parmi d’autres, avec sa seule beauté comme argument photographique. Bien vite, encouragé par Philippe GOFFART et Michel TARRIER à le rechercher activement, c’en était presque devenu une obsession. Repéré çà et là en forêt mais toujours parcimonieusement en mai/juin 2001, il me restait à en localiser le ou les dèmes à l’origine de ces spécimens épars. Il s’est avéré alors que deux colonies distinctes étaient à considérer. Disséminée en pleine forêt, une station étroite et vulnérable est opportunément liée au maintien de nombreux pieds de Succise, la plante nourricière de la chenille localement monophage. Les clairières favorables à E. aurinia sont reliées entre elles par de larges allées faisant office de ponts à l’usage de la mixité de ces maigres effectifs. La révélation de ce nécessaire va et vient entre ces réservoirs génétiques nous a incité, mon fils et moi, à entamer des travaux d’aménagements et d’élargissement des layons afin d’optimiser les chances de maintien de cette population résiduelle et en perte d’espace.
Je n’ai découvert la deuxième population, qui s’est avérée très dense, qu’en mai 2003 (première observation le 4 mai). Cette colonie était pressentie dès l’automne 2002. Le résultat de la recherche des nids d’hiver par la biologiste Violaine FICHEFET, qui s’était rendue sur les lieux suite à mes informations, s’était révélé peu positif en indices : seulement quatre nids détectés, mais il est vrai recherchés fort tardivement. Une éclosion massive en mai 2003 est venue confirmer mes intuitions, en nous révélant une très belle population. Je crains cependant pour le devenir de cette prairie communale louée en « bail à ferme », qui pour l’instant n’est heureusement pacagée que par des chevaux qui semblent respecter les massifs de Succise... du moins tant qu’ils n’étaient que deux ou trois. Depuis septembre 2003, à six sur une surface trop restreinte, affamés, ils ont commencé du bout des dents à prélever les boutons floraux de la Succise faute d’autre nourriture, ce qui n’était encore jamais arrivé jusqu’ici. Tous les plants ont de plus été fortement piétinés. Le printemps 2004 nous dira si les chenilles n’ont pas trop souffert de cette disette chevaline.(*) Des densités de 0,1 à 0,2 UGB/ha (Unités de gros bétail à l'hectare) semblent constituer le maximum tolérable, si l'on ne veut pas occasionner de dommages aux populations en place (GOFFART, 2003, communication personnelle), et ici, cette année, nous avions deux chevaux à l’hectare ! Les conséquences pourraient se révéler dramatiques pour la survie du papillon. Toute prairie livrée à un agriculteur reste toujours très vulnérable, et peut « mourir écologiquement » du jour au lendemain par le moindre amendement agressif, ou un quelconque bouleversement agro-pastoral, ce qui serait évidemment très dommageable puisque Euphydryas aurinia est une espèce protégée par la Convention de Berne et la directive « Habitats » de la CE. Classés « Natura 2000 », ces prés humides devraient au plus vite faire l’objet d’une protection intégrale pour sauver in extremis l’une des dix ou douze dernières populations wallonnes du Damier de la Succise. (Aux dernières nouvelles du début 2004, la Division Nature et Forêts par l’entremise de son Ingénieur chef de Cantonnement Jean-Pierre SCOHY, reprendra la gestion de cette prairie, en accord avec la commune de Doische. Nos appels au secours ont donc étés bien heureusement entendus). Les autres Nymphalidae Nymphalinae sont inventoriés dans le tableau qui suit.
(* ) d’après nos observations de mai 2004, la population d’Euphydryas aurinia semble très dense. Le surpaturâge n’a donc pas eu d’effet négatif sur le papillon. La population est cependant en net déclin par suite de trois années consécutives à « printemps pourris ». 2008 sera un test important pour la pérennité de l’espèce.
NYMPHALIDAE Satyrinae
Huit Satyrinae hantent assidûment le site et un neuvième ne devrait correspondre qu’à un signalement occasionnel.
Si Aphantopus hyperantus, et Maniola jurtina volent par centaines (voire par myriades pour le Myrtil), si Pyronia tithonus est toujours d’un effectif très dense, si Lasiommata megera, Melanargia galathea, Pararge aegeria, Coenonympha arcania et C. pamphilus sont ici des espèces habituelles, voire presque des vétilles, je n’ai rencontré par contre qu’un seul exemplaire d’Erebia medusa le 25 mai 2003. L’habitat ne lui convient pas et j’ai très certainement eu affaire à un sujet divagant, dispersal provenant des coteaux de Meuse qui à vol d’oiseau ne se trouvent qu’à deux à trois kilomètres du site étudié.
LYCAENIDAE
Treize Lycaenidae ont à ce jour été retrouvés.
Cinq Theclas, avec C. rubi, N. quercus, .S. ilicis, S. pruni et T. betulae.
Cinq Polyommatinae avec C. argiolus, C. semiargus, P. icarus, L. coridon et A. agestis.
Deux Lycaeninae, avec Lycaena phlaeas, et Lycaena (Heodes) tityrus.
En début mai 2004, le seul Riodininae présent en Wallonie,était enfin recensé sur le territoire avec Haemearis lucina.
Satyrium pruni s’est montré très abondant en juin 2003 avec une éclosion massive le 7 juin. Une cinquantaine d’imagos butinaient les fleurs des massifs de ronces présents dans une de nos clairières forestières.
Les adultes de Satyrium ilicis qui ne dédaignent pas plus que Satyrium pruni les ronciers en fleurs s’observent encore assez facilement. S. ilicis éclot lorsque S. pruni commence à s’éteindre, et le chevauchement ne dépasse pas huit à dix jours (première apparition le 29 juin 03).
Neozephirus quercus descend moins souvent butiner que les deux premiers frondicoles cités, ce qui fait que son abondance passe souvent inaperçue.
Les deux seuls Cuivrés repérés à ce jour n’ont pas la même fréquence.
Lycaena phlaeas, (le Cuivré commun), habituellement ubiquiste, est plutôt « rare » sur le site, excepté en 2003 ou sa présence était plus repérable.
Le Cuivré fuligineux (L. tityrus) plus répandu fréquente les lisières de forêt où il est aisé de le découvrir en juillet butinant sur l’Origan ou sur la Salicaire commune (des individus épars se rencontrent dans toute la forêt).
Quant aux Polyommatinae, si P. icarus est présent partout (sauf en forêt fermée), ce n’est jamais en grand nombre, sans doute victime de l’écocide à la gyrobroyeuse (suite futile de « Massacre à la tronçonneuse ... ») qui détruit systématiquement ses pontes en voulant assurer de beaux chemins policés, nets et propres (?), aux bas-côtés arasés. Les bermes de route et de chemins communaux étant donc systématiquement gyro-saccagés deux fois l’an minimum, seules les pontes un peu excentrées échappent ainsi à la faux moderne, aveugle et dévastatrice (j’en connais une grosse population à un kilomètre à vol d’oiseau de la propriété dans une grande mise à blanc de quelques hectares à l’abri de la gyrobroyeuse, ce qui conforte mon hypothèse concernant sa paucité dans le pourtour de la Haie Gabaux).
C. semiargus qui me semblait exceptionnel avant 2002 a été aperçu plus fréquemment en 2003 en lisière ou dans les allées très larges aux bermes fleuries de trèfles, volant tant au-dessus des prairies marécageuses (fréquente tendance hygrophile), que des prairies plus sèches, mais jamais très loin de sa plante-hôte.
Même phénomène pour Aricia agestis photographié une seule fois sur le site en 2002 puis repéré à maintes reprises en 2003.
L. coridon est recensé chaque année, mais n’est pas dans son habitat idoine. Comme pour Erebia medusa, la proximité de son berceau que sont les coteaux de Meuse explique sa sporadicité locale.
J’ai malgré tout repéré quelques individus tant mâles que femelles très frais et semblant nés sur le site, et ce, toujours le long des chemins empierrés, favorables à quelques calcicoles.
Fin août 2003, je prenais un cliché de Celastrina argiolus, portant à cinq le nombre de Polymmatinae recensés.
ESPECES POTENTIELLES
Plusieurs Rhopalocères potentiels manquent encore à cet inventaire et je continuerai à les rechercher activement.
L’absent le plus flagrant était jusqu’en 2004 Clossiana euphrosyne que je n’avais pas encore pu différencier « au vol » de Clossiana selene. Certainement observé à plusieurs reprises sans le savoir, mon seul moyen de capture étant un objectif macro, et l’ayant même probablement photographié le 11 mai 2003, mais sans avoir eu la possibilité d’observer le revers (et donc sans certitude aucune) il avait échappé à ma sagacité. J’ai dû m’autoriser quelques prélèvements traditionnels sans doute plus compatibles avec la science entomologique qu’un simple objectif macro pour l’ajouter en mai 2004 à la liste des Rhopalocères de Doische/Trou des Gattes/Haie Gabaux. Capturer/relâcher comme le font tous les observateurs aguerris est parfois bien nécessaire! Depuis 2004, il s’observe chaque année.
« Vieux fantôme », Limenitis populi serait présent sur le site (fréquentes notations personnelles avant 1980 et cité par Christophe Bauffe en 1994). Je n’ai pas personnellement réussi à le repérer récemment. Souvent victime de sa propension à « butiner » le bitume chauffé par le soleil en fin de journée, il a toujours payé un lourd tribut à la circulation automobile. Comme son repaire électif est la cime des arbres, il n’est pas d’approche aisée et passera sans doute inaperçu tant que je n’aurai pas recours à un véritable outil d’exploration de la canopée fagnarde ! Sa période de vol assez courte complique encore la difficulté de sa découverte.
Son cousin, Limenitis camilla est lui par contre assez bien représenté, volant en nombre certaines années. Et bizarrement 2003, année exceptionnelle pour bien des Rhopalocères, ne lui a pas été spécialement favorable, du moins à Doische.(en 2004, il volait en abondance, du moins les rares jours ensoleillés !)
Autres présences pressenties (ou espérées) :Plebeius argus, Satyrium w-album, Plebeius argus, voire Cupido minimus, ou encore Lycaena hippothoe… (on peut toujours rêver!)
On ne peut jamais vraiment « prouver » l’absence d’une chose... La rencontre avec un insecte reste toujours sujette aux aléas climatiques, ainsi qu’à l’état de disponibilité de l’observateur, même sur un espace restreint. Seule la fréquence des visites sur plusieurs saisons risque de réduire cet aspect aléatoire.
LES RHOPALOCÈRES « INTRUS » DE DOISCHE, OU LE DILEMME D’UN CORTÈGE CONTRASTÉ.
Dans mes relevés d’inventaire des Lépidoptères rhopalocères et hespéridés du Bois des Fagnes / Trou des Gattes / Haie Gabaux, habitat caractéristique de Fagne acide, j’ai souvent été étonné de recenser quelques présences d’espèces estimées comme calcicoles dont certaines propres aux pelouses rases.
Je me réfère ici à l’observation d’Iphiclides podalirius, de Colias alfacariensis, Polyommatus (Lysandra) coridon, Aricia agestis, Erebia medusa, toutes espèces apparemment inopportunes sur le site étudié. Il existe des papillons à double identité écologique, produisant même des morphes distinctes et induites par la nature écologique ou géologique de chaque habitat respectif. Mais ce n’est pas apparemment le cas des « intrus » rencontrés et syntopiques avec les habitants inhérents au site.
Certes, j’ai souvent vu errer ces figurants non attendus sur les rares chemins empierrés de longue date en roches calcaires, ou la flore est d’ailleurs franchement devenue calcicole, mais est-ce suffisant pour les y croire implantés ? J’ai pu contacter P. coridon ou A. agestis fraîchement éclos, mais leur présence en nombre sur les tiennes* calcaires de Foisches, à quelques petits kilomètres à vol de papillon, explique peut-être la fraîcheur de ces éventuels vagabonds.
Le cas d’Erebia medusa est le plus énigmatique. Photographié une première fois en pleine forêt un jour de pluie, il était évident que cet exemplaire réfugié dans une clairière pour se mettre à l’abri des intempéries, n’était qu’un « intrus » volant au gré des vents. Sa présence se confirme depuis lors quasi chaque année.
Les tiennes de Foisches étaient une fois de plus le berceau d’origine de cette espèce calcicole dont les stations belges sont d’ailleurs assez peu nombreuses.
Si je prends au pied de la lettre la carte géologique 58/1-2 publiée par la Direction Générale des Ressources naturelles et de l’Environnement, la partie sud de la propriété au lieu dit « Au Crestia » se trouve sur une frange d’une centaine de mètres sur sol « Frasnien » et non plus « Famennien » - Formation des Valissettes : schistes verts localement à nodules de calcaire - , d’où l’on passe progressivement en quelques centaines de mètres à la formation calcaire d’argiles noire et de schistes (Neuvillien) à celle de calcaires grossiers gris clairs du Massif de Philippeville, pour se terminer sur l’autre rive de la Meuse aux affleurements fossiles de récifs coralliens du Givetien (360.000.000 d’années). Le berceau calcaire n’est donc pas très éloigné.
La seule barrière à la dispersion des populations de lépidoptères calcicoles de Foisches, est l’imposant « désert agraire » que représente la zone réservée, ou mieux accaparée, par l’agriculture intensive, qui à réduit les « Hauts de Doische » - naguère si riches - en un milieu abiotique, dépourvu du moindre petit réservoir génétique. Il faut passer la frontière française toute proche, et circuler sur les chemins et les tiennes du village de Foisches - 3 km à vol de papillon - pour retrouver ce que les « Hauts de Doische » étaient encore dans les années 1930 d’après la littérature.
Seuls un grand volateur tel qu’Iphiclides podalirius ou un erratique aussi instable que Colias alfacariensis sont susceptibles d’atteindre par leurs propres moyens et de façon récurrente le site du Trou des gattes / Haie Gabaux. P. coridon est aussi connu pour ses capacités à prendre occasionnellement la tangente. Mais Aricia agestis est un insecte sédentaire, aux mœurs très confinées, que seul le dieu Eole serait susceptible de « projeter » là où il se cantonnera instinctivement aux quelques substrats calcaires de fortune.
Sinon, quel phénomène régit ces « intrus » ? Et verrais-je un jour Hesperia comma, Thymelicus acteon, Spialia sertorius, Cupido minimus, voire Polyommatus thersites - tous relevés à Foisches ou à Rancennes sur l’autre rive mosanne - poussés par le même vent bienfaiteur d’un inespéré repeuplement ?
« Autant en importe le vent », comme dirait plaisamment mon ami Michel Tarrier qui estime « qu'on peut davantage compter sur le vent que sur les pouvoirs publics dans l'objectif de repeupler aléatoirement un habitat » s’il n’y avait ce grand, immense et triste « mais ! » .
Car, il y a un « mais », cette année encore, ces beaux chemins de campagne, où ne passent que quelques rares vaches et tracteurs, ont de nouveau été gyrobroyés, arasés, « nettoyés » par les services communaux, détruisant en un instant, ce que la nature par le vent tente de reconstruire. Adieu Coridon, Agestis, Icarus, Phlaeas et autres habitants de ces bermes fleuries.
Messieurs les édiles communaux, Monsieur l’Echevin de l’environnement, si vous me lisez – vous avez eu la gentillesse de me reprendre en lien sur le site officiel de Doische - expliquez moi pourquoi, vous préférez des bords de chemins ras, nets et semblant « propres », mais « morts », complètement abiotique, à ces bermes fleuries, pleines de vie ? Sont-ce ces herbes « folles » qui vous hérissent ? La notion de « Fauchage tardif » est pourtant ancienne, pourquoi n’est-elle JAMAIS appliquée ? Pourquoi faut-il macadamiser le Ravel**, pourquoi en gyrosaccager les abords trois à quatre fois l’an ? Bétonnez tout, et qu’on en finisse une fois pour toutes !
Les Bleus nacrés , les Argus bruns , les Orvets, les Lézards, les Couleuvres, et les fleurs sauvages vous demandent un tant soit peu de respect de l’aire de leurs amours, jeux, vie et mort.
Pitié pour eux, pitié pour nous, et que ce petit aparté déclenche une réflexion salutaire.
La Terre en a besoin, nous en avons besoin. Merci.
· * Collines couvertes de terrains incultes, friches, pelouses calcaires typiques de la Calestienne.
· ** Le Ravel est un réseau de voies récréatives, élaborées par la Région Wallonne et réservées aux piétons, cyclistes et cavaliers, empruntant les anciennes voies ferrées désaffectées, anciennement biotope de première importance pour quantité de Lycènes qui y trouvaient une flore adaptée. Actuellement lieu quelque peu « aseptisés » par la pose d’infrastructures en « dur » aux abords écorchés par un gyrobroyage répétitif.
NOTATIONS DE PRÉSENCES ANCIENNES RELEVÉES DANS LA LITTÉRATURE.
Melitaea cinxia a été signalé de Doische (sans précisions) en 1934 de même qu’à Romedenne, comme abondant. (cf. DERENNE, F., 1935, Remarques sur la faune belge. Lambillionea, 35 ème année (1) :18.
En 1970 (cf. FONTAINE, M. & HECQ, J. 1971, Principales captures de Lépidoptères et observations de l’année 1970. Lambillionea, LXXI (3-4) :33),
M. cinxia n’a pas été noté dans les stations connues de l’Entre-Sambre-et-Meuse !! Pourtant, plus d’une trentaine de stations étaient connues dans le district calcaire-mosan (prov. Namur et partiellement Luxembourg). Une disparition continuelle des populations s’y est produite pendant les années 1960.
VERSTRAETEN, Ch., GASPAR, Ch, LAGRANGE, B. & ANCELOT, Ph., 1986, Évolution des populations de Lépidoptères diurnes en Belgique de 1960 à 1983. Bull. Annls Soc. r. belge Ent.. 122 (1986) : 55-79, confirment de façon dramatique ce que FONTAINE & HECQ (1971) ont constatés.
M. cinxia ne s’observe plus que très épisodiquement depuis 1975 au sud du sillon Sambre et Meuse. Il a cependant encore été photographié en 2003 par Stéphane Claerebout à la Montagne aux Buis à Nîsmes.
Dans leur document de travail sur la répartition des rhopalocères de la province de Luxembourg, P. & M. TAYMANS, 1999 in Bull. Cercle Lépidoptéristes Belgique, XXVIII (1), écrivent que M. cinxia aurait disparu de la Famenne et des Ardennes, phénomène de raréfaction qui aurait débuté pendant les années 1950.
Encore franchement commune en Lorraine belge pendant les années 1980, M. cinxia semble s’y maintenir çà et là, ce que j’ai pu observer personnellement en juin 2004.
GOFFART, Ph., & DE BAST, B., 2000, dans leur atlas préliminaire des papillons de jour en Wallonie. Publication du Groupe de Travail Lépidoptères, Thomas & Chabot, Marche, ne connaissent plus que 11 populations de M. cinxia, toutes situées en Lorraine belge mais sans donner de précisions.
Lyceana (Paleochrysophanus) hippothoe (LINNÉ, 1757) a été signalé de Doische (sans précisions) en 1984 (cf. ALEXIS, R., 1985, Première contribution au recensement des Lépidoptères Rhopalocères de la région de Mariembourg et environs. Observations et captures de l’année 1984. Lambillionea LXXXIV (9-10) : 95.
Maculinea rebeli [HIRSCHKE] aurait été capturé à Doische (sans précisions) par J. DRUET dans les années 1930 (cf. VIGNOUL, H., 1934, Où devons nous chasser ? Lambillionea, 34 ème année, n°3 : 67).
À cette époque, la séparation M. rebeli de M. alcon n’avait pas encore été faite et BERGER, L., 1946, Maculinea rebeli HIRSCHKE, bonna species, Lambillionea XLVI (6-10) : 95-110, pl. 1, 9 figs), regrette de ne pas avoir pu examiner les captures de DRUET faites dans l’Entre-Sambre-et-Meuse afin de statuer sur leurs populations . Il s’est avéré par la suite que M. alcon est confirmé dans la Campine anversoise et limbourgeoise où l’espèce fréquent les landes à bruyères humides hébergeant sa plante hôte, Gentiana pneumonanthe.
Aucune donnée sur M. alcon dans l’Entre-Sambre-et-Meuse ne nous est connue à l’heure actuelle, alors que Gentiana pneumonanthe y a été trouvée. Par contre, M. rebeli était connu de la Région mais probablement disparu sauf dans le secteur français de Givet – Chooz – Fromelennes, où il s’est raréfié considérablement depuis les années 1990.
Inventaire documenté des Rhopalocères de Wallonie, présents sur le territoire de Doische – Trou des Gattes – Haie Gabaux, classés par catégories de menaces selon les critères de l’UICN
Jean DELACRE (2001/2004)
En danger critique
(Critically endangered)
Nymphalidae
Euphydryas aurinia
Le Damier de la Succise est abondant en prairie, plus rare et localisé en forêt.
Univoltin (V-VI)
Sylvicole et praticole
Clairières, prairies humides ou sèches à Succisa pratense.
Nymphalis antiopa
Le Morio est très rare, voire en grand danger d’extinction.
Une seule génération qui estive et hiberne (double diapause), pouvant se rencontrer en avril, juillet/août, et parfois en automne
Sylvicole.
Limenitis populi
Le Grand Sylvain est très rare.
À retrouver (dernière observation en 1994).
Univoltin (VI – VII)
Sylvicole et frondicole.
En danger
(Endangered)
Papilionidae
Iphiclides podalirius
La présence du Flambé est accidentelle.
Univoltin (V-VI), rarement seconde génération (VIII)
Versants xériques à Prunus spinosa. Se reproduit en adrets.
Volateur.
Pieridae
Colias alfacariensis
Sa présence est accidentelle.
Bi à trivoltin (IV à X)
Calcicole, pelouses sèches.
Nymphalidae
Mellicta athalia
Le Damier Athalie est dispersé en petites populations éparses, parfois denses.
Univoltin (V-VI)
Bois clairs à Melampyrum pratense, lisières, clairières, pelouses sèches.
Clossiana euphrosyne
Localement bien présent
Univoltin (III-VII)
Lisières et clairières ensoleillées, chemins forestiers.
Clossiana dia
Repéré à une seule reprise en août 2004.
Visible maintenant chaque année, depuis les travaux d’aménagement.
Bi à trivoltin (IV-IX)
Pelouses calcaires, lisières forestières chaudes et pas trop sèches, endroits broussailleux non loin des zones boisées.
Vunérable
(Vulnerable)
Hesperiidae
Carcharodus alceae
La Grisette est très peu abondante. Quelques rares observations sur des talus secs.
Plurivoltin (IV à X)
Pelouses sèches, friches.
Carterocephalus palaemon L’Echiquier est abondant sur quelques modestes surfaces de clairières ensoleillées.
Univoltin ((V à VI)
Lisières, clairières humides, plutôt hygrophile.
Lycaenidae
Hamearis lucina
Localement peu présente, et plutôt accidentelle bien que sa plante-hôte y soit commune.
Univoltin (III-VII)
Rarement deux générations les années de fort ensoleillement.
Lisières, clairières, layons forestiers.
Aricia agestis
Le Collier-de-corail est plutôt rare sur quelques bermes de chemins empierrés, mais se rencontre chaque année.
Bi à trivoltin (IV-X)
Prairies maigres, pelouses sèches, lisières et bois clairs.
Cyaniris semiargus
Le Demi-Argus vole en forêt sur des chemins largement ouverts, aux bermes recouvertes de trèfles. Toujours peu répandu.
Bivoltin (IV à VI et VII à X)
Praticole, champs de trèfles, prairies fleuries.
Lycaena tityrus
Le Cuivré fuligineux est répandu sur toute l’aire, en lisière comme en forêt.
Jamais abondant, mais toujours bien présent.
Plurivoltin (V à X)
Mésophile, évite les biotopes trop secs.
Lysandra coridon
L’Argus bleu-nacré est limité aux chemins empierrés, plutôt accidentel en provenance des coteaux de Meuse.
Univoltin (VII à IX)
Calcicole, prairies maigres.
Satyrium ilicis
La Thecla de l’yeuse est abondante sur certains ronciers en lisières, ou dans les chemins très dégagés.
Univoltin : (VI à VIII)
Haies, lisières et bois clairs.
L’imago aime butiner les fleurs de ronce.
Satyrium pruni
La Thécla du prunier est assez commune. Eclosions parfois massives en juin.
Univoltin (V à VII)
Aime les lisières, les clairières et les bois clairs.
Nymphalidae Nymphalinae
Apatura ilia
Le Petit Mars changeant est abondant en forêt, principalement sous la forme clytie.
Univoltin (VI – VII)
Sylvicole et frondicole.
Argynnis adippe
Le Moyen Nacré est très localisé en clairières et peu abondant.
Univoltin (VI à VIII)
Prairies bocagères, landes, clairières ensoleillées.
Argynnis aglaia
Le Grand Nacré est localisé en clairières forestières
Univoltin (VI à VIII)
Lisières, clairières fleuries.
Clossiana selene
Le Petit Collier argenté est très abondant dans les chemins forestiers et les clairières bien ouvertes.
Bivoltin (V à VII et VII à VIII)
Préférences pour les biotopes sur sol acide, mais assez éclectique.
Melitaea diamina
Localement : la Mélitée noirâtre est assez abondante dans certaines clairières.
Univoltin (V à VIII)
Clairières, tourbières, bois clairs plutôt humides.
Nymphalis polychloros
La Grande Tortue est bien présente, mais plutôt rare. (en nombre cependant au printemps 2004)
Univoltin, mais estive et hiberne.(III – VI à VIII – parfois IX à X)
Bois clairs et lisières.
Nymphalidae Satyrinae
Coenonympha arcania
Le Céphale est abondant sur quelques chemins secs.
Univoltin (V à VII) et parfois deuxième génération partielle de (VIII à X)
Bois clairs, lisières, prairies bocagères.
Erebia medusa
Le Moiré franconien est accidentel (en provenance des coteaux de Meuse)
Univoltin (V à VII)
Pelouses sèches.
Menacé
(Threatened)
Issoria lathonia
Le Petit Nacré est rare, accidentel et vagabond.
Deux générations possible.
(III à X)
Praticole, friches, prairies fleuries.
Faible risques
(Lower risk)
Hesperiidae
Erynnis tages
Le Point-de-Hongrie est très répandu dans les clairières dégagées, et sur les bermes des larges layons.
Bivoltin (IV à VIII)
Prairies maigres, clairières florifères, bermes de routes.
Pyrgus malvae
L’Hespérie de la mauve est présente partout et assez abondante certaines années.
Univoltin à confirmer (IV à VI)
Clairières, layons forestiers, prairies rases.
Pieridae
Aporia crataegi
Le Gazé est très abondant dans les chemins aérés, les friches et les clairières
Univoltin : (V à VII)
Bois clairs, haies, lisières, prairies bocagères.
Leptidea sinapis (*)
La Piéride de la moutarde est très abondante toute l’année de mars à septembre.
Bivoltin : (IV à VIII)
Clairières, lisières, prairies embroussaillées.
Lycaenidae
Callophrys rubi
La Thécla de la ronce est peu répandue.
Univoltin (IV à VI)
Lisières et clairières, landes, prairies recolonisées par la broussaille.
Thecla betulae
La Thecla de bouleau est probablement plus abondante qu’il n’y paraît.
Univoltin (VII à X)
Bois clairs, lisières, haies.
Nymphalidae Nymphalinae
Apatura iris
Le Grand Mars changeant est abondant sur quelques aires de vol.
Univoltin (VI à VII)
Clairières en forêt avec présence de saules marsaults.
Sylvicole et frondicole.
Argynnis paphia
Localement : le Tabac d’Espagne est très abondant chaque année.
Univoltin : (VI à IX)
Clairières florifères, lisières forestières, bois clairs.
Brenthis ino
Le Nacré de la sanguisorbe est très abondant dans certaines friches, et dans les layons humides mais ensoleillés.
Univoltin : (V à VII)
Clairières humides, friches. Plutôt hygrophile.
Limenitis camilla
Le Petit Sylvain est très abondant certaines années. 2003 fut une année moyenne.
Univoltin : (VI à VIII)
Clairières forestières, lisières, layons ensoleillés.
Sylvicole et frondicole.
Nymphalidae Satyrinae
Melanargia galathea
Le Demi-deuil est très abondant.
Univoltin : V à VIII)
Ubiquiste.
À la limite d’être menacé
(Near threatened)
Papilionidae
Papilio machaon
Le Machaon se reproduit sur le site. Il n’est jamais abondant, mais régulier. Observation d’une ponte sur la reine des prés, ce qui semble peu habituel.
Bivoltin (IV à IX)
Milieux ouverts.
Volateur.
Non menacé
(Safe)
Hesperiidae
Ochlodes venatus
La Sylvaine est très répandue.
Univoltin : (VI à IX)
Clairières humides, lisières bocagères, chemins forestiers.
Thymelicus lineolus
L’Hespérie du dactyle est très abondante.
Univoltin : (VI à VIII)
Clairières, prairies embroussaillées, milieux ouverts.
Thymelicus sylvestris
L’Hespérie de la houque est très abondante.
Univoltin : (VI à VIII)
Clairières forestières, prairies sylvatiques, milieux ouverts.
Pieridae
Anthocharis cardamines
L’Aurore est très répandue au printemps, le mâle patrouillant sur les lisières en se posant peu.
Univoltin (IV à VI)
Clairières, lisières forestières, prairies maigres.
Gonepteryx rhamni
Le Citron est le plus abondant de tous les papillons de Doische avec M. jurtina. La bourdaine omniprésente explique ce festival papillonnant de jaune.
Univoltin (VI à X et II à V)
Sort d’hibernation très tôt dés les premiers rayons de soleil (février)
Clairières florifères, lisières, allées dégagées.
Pieris brassicae
La Piéride du choux vole partout, mais en petit nombre.
(IV à X) en deux à quatre générations
Présente des jardins à la clairière en forêt.
Pieris napi
La Piéride du navet est très abondante.
(III à X) en deux générations
Ubiquiste, milieux ouverts, ripisylves, et bois clairs.
Pieris rapae
La Piéride de la rave est très abondante.
(III à X) en 3 à 4 générations
Milieux ouverts variés.
Lycaenidae
Celastrina argiolus
L’Argus à bandes noires semble peu abondant.
Bivoltin à trivoltin les années fastes (III à VI & VI à VIII) (parfois IX à X)
Lisières, haies, clairières. Ne quitte pas le strate arbustive.
Lycaena phlaeas
Le Cuivré commun était plutôt rare en 2001 et 2002. Plus présent en 2003, et même abondant en septembre.
Plurivoltin : (IV à X)
Lieux ouverts, friches fleuries et ensoleillées.
Neozephyrus quercus
La Thécla du chêne est abondante mais difficilement observable sans jumelles, et sans lever les yeux.
Univoltin : (VII à VIII)
Lisières de chênaies, chemins forestiers avec chênes.
Polyommatus icarus
L’Argus bleu est répandu, mais paie un lourd tribut à la gyrobroyeuse.
Plurivoltin (IV à X)
Landes, clairières forestières, prairies florifères, bords de chemins herbeux.
Nymphalidae Nymphalinae
Aglais urticae
La Petite Tortue était rare en 2001, en profusion en 2003. Fluctuations très importantes des effectifs.
Souvent bivoltin (III à X) (hiverne parfois)
Ubiquiste, milieux ouverts.
Araschnia levana
La Carte géographique est abondante sur les pourtours de la forêt, plus rare en forêt profonde.
Bivoltin (IV à IX)
Lisières buissonneuses, humides et ensoleillées et bois clairs.
Inachis io
Le Paon-du-jour est très commun. Aime butiner sur l’Eupatoire à feuilles de chanvre
Bivoltin (III à X) (hiverne souvent)
Lieux ouverts, et fleuris, y compris en forêt.
Polygonia-c-album
Le Robert-le-diable est assez commun.
Bivoltin (III à VIII) (hiverne souvent)
Bois clairs, lisières forestières, haies, prairies maigres de bordures forestières.
Nymphalidae Satyrinae
Aphantopus hyperantus
Localement : le Tristan est très abondant dans les chemins forestiers, et sur les prairies maigres de bordure.
Univoltin : (VI à VIII)
Plutôt hygrophile, aimant les bois clairs, les landes et prairies humides.
Coenonympha pamphilus
Le Procris (Fadet commun) est parmi les premiers à être observé dés la fin mars. Abondant.
Plurivoltin (IV à X)
Friches, prairies de pourtour de forêt, lisières.
Lasiommata megera
La Mégère se cantonne sur les chemins empierrés secs. Se rencontre plus rarement en forêt.
Plurivoltin : (IV à X)
Chemins secs et rocailleux.
Maniola jurtina
Le Myrtil est le plus abondant de nos papillons (avec G.rhamni).
Semble partiellement estiver, pour réapparaître en août.
Univoltin : (VI à X)
Ubiquiste.
Entre en quiescence lors des grandes chaleurs.
Pararge aegeria
Le Tircis est abondant dans les layons ombragés.
(IV à X) en deux à trois générations
Bois, lisières et parcs.
Comportement territorial très marqué.
Sciaphile.
Pyronia tithonus
L’Amaryllis vole partout en forêt et en lisières, très abondant..
Univoltin : VII à IX)
Clairières, lisières, prairies embroussaillées.
Visiteurs / Migrateurs
(Vagrant / Migrant)
Pieridae
Colias crocea
Le Souci est peu abondant sauf en 2003, ou sa présence était journalière de fin juillet à août, et très abondant en septembre.
Bi à trivoltin (IV à X)
Milieux ouverts et aérés.
Nymphalidae
Vanessa atalanta
Le Vulcain est très abondant.
(Abondance variable d’une année à l’autre).
Migrateur se reproduisant localement en été (III à XI) (hiverne parfois)
Milieux ouverts, lisières et bois clairs.
Cynthia cardui
La Belle-Dame est très abondante. (Abondance variable d’une année à l’autre).
Migrateur se reproduisant en été (III à XI)
Ubiquiste en milieux ouverts.
(*) Leptidea reali n’a pas encore été découvert dans la région, quoi qu’il soit présent dans le Virtonnais.
BIOINDICATION, BIOSURVEILLANCE ?
Agents essentiels des cycles biologiques, réagissant ipso-facto au moindre effet nocif (notamment au niveau des plantes-hôtes dont ils sont tributaires), par un recul ou une extinction, les papillons sont les véritables révélateurs pour le diagnostic d’une telle situation. Solidaires de chaque écosystème, ils s’en avèrent être les meilleurs marqueurs synécologiques. Leur influence sur les écosystèmes se manifeste autant par leur présence que par leur absence. En ce sens, les espèces les plus signifiantes ne sont pas à considérer individuellement, mais collectivement, un peu sur le mode d’une guilde. (sensu Tarrier, 2004, comm.pers.).
À Doische, il y a 80 ans volaient de concert dans les mêmes prairies humides Melitaea cinxia, Euphydryas aurinia et Lycaena hippothoe. En 2004, seul Euphydryas aurinia à réussi à survivre tant bien que mal et très ponctuellement. Nous ne leurrons donc pas, ce type de biotope est bien malade, sa dégradation progressive étant arrivé à un stade paroxystique et quasi irréversible.
La plupart de ces « papillons marqueurs » sont monophages ou oligophages, et étroitement inféodés à des plantes-hôtes sensibles et vulnérables. Il s’agit donc d’une panoplie d’éminents indicateurs biologiques qui réagissent aux modifications nocives par un recul, puis par la disparition. Les « insectes-outils » sont censément moins maniables mais sans nul doute plus précis que les vertébrés ou les plantes, tant pour la gestion et la sélection des sites à protéger, que pour l’évaluation de l’incidence biologique en baisse des surfaces menacées, en un mot pour la conservation du patrimoine naturel au service des populations rurales fragilisées par de nouvelles donnes économiques. Les espèces parfaitement sténoèces, hautement vulnérables, ne supportant pas un équilibre rompu par la moindre intervention, pression ou nuisance, ces dernières sont des bioindicatrices emblématiques de la valeur d’un milieu, aussi nommées « espèces-ombrelles » ou « espèces clé-de-voûte » et veillent à la naturalité de l’habitat.
L’utilisation de données entomologiques pour une gestion à long terme exige une validation continue des dites données. Les espèces d’insectes, dans leur grande majorité, ne sont identifiables que sous la loupe binoculaire, tandis que leur récolte sur le terrain nécessite des méthodes de prospection et d’échantillonnage adaptées. Chaque donnée unitaire implique donc : suivi de visites, capture, identification, archivage des données, et suivi dans le temps.
BIBLIOGRAPHIE
BAGUETTE, D. & al., 1992,
Modification de la distribution et du statut des lépidoptères Rhopalocères de Belgique depuis 1990.
Mém. Soc. Roy. Belge Ent. 35 :591-596.
BAILEY, K.E.J, PORTER, K., & al.,
The moths and butterflies of Great Britain, vol. 7(1) :234-237 (E. aurinia) édit. MAITLAND EMMET, A. & HEATH, J., HARLEY Books Colchester, U.K.
BARNETT, L.K. & WARREN M.S., 1995,
Species Action plan : Marsh Frittilary : Eurodryas aurinia.
Butterfly conservation, Dorset.
BINK, F.A.,1992,
Ecologische Atlas van de Dagvlinders van Noordwest-Europa.
Schuyt & Co, édit.,Haarlem, Pays-Bas : 512 p. (E. aurinia : 392-393) nb. Pl. coul. et cartes de répart.)
CAVELIER, E., RENNESON, J.L., TAYMANS, P. & VALENNE, Y.,1998,
Les papillons de Lorraine belge. Synthèse et cartographie de cinq années d’études des papillons de la Lorraine belge.
Rhoplalocera 1990-1994. Notes Faunistiques de Gembloux N° 34 : 3-106 (E. aurinia : 60,98,101).
CHINERY, M. & LERAUT, P., 1998,
Photo-guide des Papillons d’Europe.
Delachaux et Niestlé – Paris.
DESCIMON, H., ZIMMERMANN, M., COSSON, E., BARASCUD, B. & NÈVE, G.,
Diversité génétique, variations géographiques et flux génétiques chez quelques lépidoptères rhoplalocères français.
Génét.Sel. Evol. 33 (Suppl.1) INRA, EDP Sciences : 223-249 (E. aurinia : 226, 236-245).
EBERT, G., 1991,
Die Schmetterlinge Baden-Würtemberg, Band 1 : Tagfalter 1
E. Ulmer Verlag, Stuttgart : 552 p. nbr. Pl. coul. et cartes de répart. (E. aurinia : 542-549).
FICHEFET, V., BAUGNÉE, J-Y. & GOFFART, P.,
"Nouvelles lépidoptérologiques" (7).
Sur internet:
http://mrw.wallonie.be/dgrne/sibw/especes/ecologie/papillons/ISB_SURWAL/News Lycaena/Lycaenanews7.htm
FISCHER, K., 1997,
Zur Ökologie des Skabiosen-Schecken Falters Euphydryas aurinia [ROTTEMBURG, 1775] (Lepidoptera : Nymphalidae).
Nachr. Entomol. Ver. Apollo, Frankt. a.M., N.F., 18 :287-300.
FITTER, R., FITTER, A., BLAMEY, M., 1996,
Guide des fleurs sauvages.
Delachaux et Niestlé, Paris.
FONTAINE, M., LEESTMANS, R., DUVIGNEAUD, J., 1983,
Les Lépidoptères de la partie méridionale de l’Entre-Sambre-et-Meuse et de la pointe de Givet.
Linneana Belgica, Pars IX 1983- N° 1 mars 1983- Volume jubilaire.
FORD, M.D., & FORD, E.B., 1930,
Fluctuation in numbers and its influence on variation in Melitaea aurinia ROTT.
Trans. Roy. Ent. Soc. London, 78 :345-351.
GELBRECHT, J., RICHTERT, A.& WEGNER, H., 1995,
Biotopansprüche ausgewähter von Austerben bedrohter oder verschollener Schmetterlingsarten der Mark Brandeburg (Lep.).
Ent.Nachr.u.Ber. 39 : 183-203.
GERBER, H., 1972,
Speziation und Biologie von E. aurinia aurinia und E. aurinia debilis F. glaciegenita. Mitt. Ent. Gesel. Basel, N.F., 22 :73-87.
GOFFART, P. & DE BAST, B., 2000,
Atlas préliminaire des papillons de jour de Wallonie, « Groupe de travail Lépidoptères »,
Thomas & Chabot, Marche.
GOFFART, P., BAGUETTE, M., & DE BAAST, B., 1992,
La situation des Lépidoptères Rhopalocères en Wallonie ou que sont nos papillons devenus ? Bull. Annls. Soc. Roy. Belge d’Ent. 128 :355-392.
GOFFART, P., BAGUETTE, M., DUFRÊNE, M., MOUSSON, L., NÉVE, D., SAWCHIK, J., WEISERBS, A., LEBRUN, P., 2001,
Gestion des milieux semi-naturels et restauration de populations menacées de papillons de jour. Travaux N° 25 Direction générale des ressources naturelles de l’Environnement, Division Nature et Forêts, Direction de la Nature, Jambes Belgium.
GOFFART, P., NÉVE, G., MOUSSON, L., WEISERB, A., BAGUETTE, M. & LEBRUN, P., 1996,
Situation actuelle, exigences écologiques et premiers résultats de deux tentatives de réintroduction du Damier de la Succise ( Euphydryas aurinia ), un papillon en danger d'extinction en Wallonie.
Cahiers des Réserves Naturelles-RNOB, n° 9.
HIGGINS, L. G., 1950,
A descriptive catalogue of the Palearctic Euphydryas aurinia.
Trans.Roy. Ent. Soc. Lond., 101 (12) : 437-489.
LAFRANCHIS, T., 2000,
Les Papillons de jour de France, Belgique et Luxembourg et leurs chenilles. Collection Parthénope, éditions biotope, Mèze (France).
LEWIS, O. T. & HURFORD,C., 1997
Assessing the status of the marsh frittillary butterfly (Eurodryas aurinia) : an example from Glamorgan, U.K. Journal of Insect Conservation 1 :159-166.
MAES, D. & VAN DYCK, M.,1996,
Een gedocumenteerde Rode lijst van de dagvlinders van Vlaanderen.
Mededel. Inst. Natuurbeh. (1) :154p. (E. aurinia : 84-85).
MAES, D. & VAN DYCK, M., 1999,
Dagvlinders in Vlaanderen, édit. Stichting Leefmilieu KBC :Antwerpen : 480 p., nbr. Pl. coul. et cartes de répart. (E. aurinia : 304-306,431).
MAZEL, R.,1977,
Première contribution expérimentale à la connaissance taxonomique et phylétique de quelques formes d’Eurodryas aurinia,
Entomops, N° 44 : 103-112.
MAZEL, R., 1982,
Exigences trophiques et évolution dans les genres Euphydryas et Melitaea lato sensu. Ann. Soc. Ent. Fr., (N.S.), 18(2).
MAZEL, R., 1982b,
Seconde contribution expérimentale à la connaissance taxonomique et phylétique de quelques formes d’Eurodryas aurinia [ROTTEMBURG]. Intérêt biogéographiques et phylétiques de deux nouvelles sous-espèces d’Eurodryas aurinia ROTT.(Lep. Nymphalidae),
Alexanor 12(7) : 303-316.
MAZEL, R., 1986,
Structure et évolution du peuplement d’E. aurinia [ROTTEMBURG] (Lepidoptera) dans le sud-ouest européen.
Vie et Milieu 36 (3).
MAZEL, R., 1984,
Trophisme, hybridation et spéciation chez E. aurinia [ROTT.]
Thèse. Université de Perpignan : 335 p.
« SIBW , 2001 », 2001,
TOLMAN & LEWINGTON. 1999,
Guide des Papillons d’Europe et d’Afrique du nord,
Delachaux et Niestlé, Paris.
VALENNE,Y. & TAYMANS, P., 1998,
Contribution à l’étude de la faune entomologique belge. Les Rhopalocères des zones humides en Lorraine et Ardenne belge.
Parcs et Réserves, rev.trim. Cons. Nat. Gest. dur. d’Ardenne et Gaume, vol.53 (3) : 26-30.
VERSTRAETEN, Ch., 1971,
Atlas provisoire des Insectes de Belgique, carte N° 564 (décembre 1971)
Fac. Sc. Agron. État, Zool. Gént. et Faunistique, Gembloux.
VERSTRAETEN, Ch., GASPAR, C., LAGRANGE, B. & ANSELOT, P., 1986,
Évolution des populations de Lépidoptères diurnes en Belgique de 1950 à 1983. I : Nymphalidae.
Bull. Annls Soc. Roy. Belge Ent.., 122 : 55-79 (E. aurinia : 72).
WARREN, M.S.,1986,
Notes on habitat selection and the larval host-plants of the brown argus, Aricia agestis (D.&S.), Marsh frittilary, Eurodryas aurinia [ROTTEMBURG], and Painted lady, Vanessa cardui [L.] in 1985.
Entomol. Gaz.,37 :65-67.
WARREN, M.S.,1994,
The U.K. status and suspected metapopulationstructure of threatened European butterfly, the Marsh frittillary Eurodryas aurinia.
Biol.conser., 67 :239-249.
WAYENBERGH, M., 1998,
Le parasitisme, facteur-clé de la survie des populations locales des Rhopalocères ?
Parc et Réserves, rev. trim. Cons. Nat. Gestion dur. d’Ardennes et Gaume, vol.53 (3) :8-11.
WYNHOFF, I., VAN SWAAY, C., VAN DER MADE, J., 1999,
Veldgids dagvlinders, KNNH, Utrecht.
LEXIQUE ECO-ENTOMOLOGIQUE
(Domaine franco-rhénan)
Acidophile : Qui croît ou vit sur un sol acide.
Adventice (végétation) : Qui croît dans un milieu cultivé ou géré sans avoir été semé (synonyme : plante de fourvoiement).
Alaire : Relatif à l’aile.
Allochtone : Espèce exotique introduite dans un milieu (antonyme : autochtone).
Adret : versant d’une vallée exposé au soleil (antonyme : ubac).
Amendement : Toute pratique ou substance destinée à améliorer les propriétés physiques d’une terre à des fins agricoles.
Anémophile : Ayant une préférence pour les lieux aérés ou ventés.
Anthropique : Dû à l’homme.
Anthropozoïque : Se dit des influences biotiques dépendantes de l’homme.
Aposématique : Avertissante, prémonitrice, répulsive, en parlant de la livrée bariolée des espèces toxiques ou venimeuses (exemples : guêpes, serpents-corail, zygènes…).
Aptère : Dépourvu d’ailes.
Aquicole : Qui vit dans l’eau (et dulçaquicole : qui vit dans l’eau douce).
Biocénose : Ensemble des végétaux, des animaux et des microorganismes vivant en étroites interrelations dans un biotope ou une niche écologique (partie du biotope).
Biodiversité : Ensemble des patrimoines génétiques des espèces végétales et animales vivant dans un biotope donné, et par extension, dans la totalité de la biosphère.
Biomasse : Quantité globale de matière organique dans un temps donné.
Biome : Milieu.
Biotope : Composition « non vivante » et physico-chimique de l’écosystème, hébergeant une flore et une faune déterminées.
Bivoltin (ou digoneutique) (entomologie) : Qui produit deux générations annuelles successives.
Bouquet : Groupe d’arbres d’âges ou de dimensions similaires.
Brométum : Prairie sèche constituée de bromes (graminées du genre Bromus).
Caducifolié : A feuillage caduc.
Calcicole : Qui croît ou évolue sur des substrats calcaires.
Calcifuge : Qui ne se plait pas en terrain calcaire.
Cariçaie : Prairie humide essentiellement formée de laîches (cypéracées du genre Carex).
Cépée : Ensemble de rejets issus d’une même souche.
Chablis : Arbres renversés ou abîmés par des facteurs naturels.
Chorologie : la chorologie examine la diffusion des différents ordres et espèces de végétaux ou animaux dans tous les espaces vitaux qui peuvent être colonisés. Elle analyse les causes climatologiques, pédologiques, historiques et géographiques à l'origine de la formation des habitats des espèces particulières, des vies communes ou des éléments de la végétation.
Climax : Stade final et équilibré de l’évolution naturelle de la végétation dans son milieu.
Cline : Ensemble de variations légères qu’accuse graduellement une espèce sur son aire de distribution. Certaines espèces évoluent selon un cline, d’autres pas.
Communauté : Ensemble d’individus appartenant à plusieurs espèces, vivant à un endroit donné, à un moment donné.
Couvert (forestier) : Ecran formé par l’ensemble des frondaisons d’un peuplement.
Cryptique : Ornementation destinée à soustraire l’insecte au regard de ses prédateurs.
Défens (ou défends) : Régime forestier de protection avec interdiction d’exercer les droits d’usage.
Dème : Unité populationnelle ou population.
Diagnose : Description scientifique d’une espèce ou autre taxon.
Diapause : Période de repos obligatoire régulée par des facteurs internes, correspondant à une suspension du métabolisme au cours d’une quelconque phase du cycle biologique. Ne pas confondre avec la quiescence, état torpide induit par des facteurs externes comme la chaleur.
Dimorphisme : Différence d’aspects entre les individus mâles et femelles (= dimorphisme sexuel), ou entre des sujets de générations successives (= dimorphisme saisonnier).
Dispersal : Vocable anglais et utilisé en faunistique pour qualifier un élément provenant d'une dispersion accidentelle. S'il rencontre des conditions favorables (plante-hôte ou de subsitution, éco-climat, etc.), l'élément divaguant peut alors établir une souche et l'espèce coloniser ainsi un nouveau territoire.
Drageon : Pousse aérienne née d’une racine, créant elle-même des racines.
Ecobuage : Incinération de la strate herbacée préalablement arrachée puis épandage des cendres.
Écosystème : Unité fonctionnelle de base en écologie comprenant le biotope et la biocénose associée.
Ecologie (scientifique) : Science étudiant les relations entre un être vivant et son milieu. Le chercheur correspondant est un écologue, et non un écologiste !
Ecotone : Zone de transition marquant le passage entre deux écosystèmes de nature différente (synonyme non utilisable en écologie : interface).
Edaphique : Qui se rapporte au sol.
Edaphologie : Science du sol impliquant des notions morphologiques, lithologiques et pédologiques.
Edaphon : Ensemble de la flore microbienne et de la faunule vivant dans l’eau interstitielle des sols.
Emondage : Enlèvement des branchettes gourmandes qui surgissent d’un tronc suite à une mise en lumière.
Endémique : Espèce originaire et exclusive du pays dans lequel elle vit.
Enrésinement : Opération consistant à planter une espèce résineuse.
Epigénique : Vivant à la surface du sol.
Equienne : Du même âge (pour un peuplement forestier).
Erémicole ou sub-saharien : habitant l’érémial, soit le pourtour de la steppe désertique.
Ethologie : Science du comportement des espèces dans leur milieu naturel.
Euryèce : Qui présente des exigences écologiques peu prononcées et donc apte à se développer dans les biotopes les plus variés (synonyme : ubiquiste ; antonyme : sténoèce).
Eutrophisé : Concernant le milieu aquatique, artificiellement surfertilisé par les pollutions agricoles et/ou urbaines.
Floricole : Qui visite les fleurs pour en soutirer la source nectarifère.
Fontinal : Vivant à proximité immédiate d’une source.
Fourré : Stade précoce et pratiquement impénétrable de développement du semis naturel
Frondicole : Sylvicole adepte de la frondaison (qui hante assidûment la frondaison)
Fruticée : Formation végétale composée d’arbustes et de buissons, souvent épineux.
Habitus : Aspect extérieur permettant la diagnose d’une espèce.
Héliophile : Qui affectionne le soleil et la lumière (antonyme : sciaphile).
Hôte (plante-hôte) (entomologie) : Plante nourricière dont est tributaire une espèce pour les recours trophiques de sa larve.
Hibernarium : Refuge individuel ou collectif servant à l’hibernation de la chenille et constitué d’une réseau soyeux.
Homotypie : Ressemblance d’un animal avec un objet inanimé pour en assurer un camouflage efficace (homochromie avec les couleurs, homomorphie avec les formes…).
Hydrophytes : Végétaux aquatiques nageants (potamaie formée de potamots, myriophylles, utriculaires…) et flottants (nupharaie pour les nénuphars, nymphéas, renouée amphibie…).
Hygrophile : Tributaire de l’humidité.
Imago : Insecte à l’état sexué (synonyme : adulte).
Lisier : Engrais naturel et liquide constitué d’excréments et d’urines des animaux domestiques (bovins, porcins, ovins), avec fort peu de litière.
Livrée : Aspect extérieur, robe d’un animal.
Mégaphorbiée : Association végétale composée d’arbustes, de plantes à grandes feuilles et à port élevé.
Mésofaune : Animaux de taille moyenne peuplant le sol : hexapodes (insectes et entognathes), acariens, annélides, mollusques…
Mésophile : Qui affectionne un milieu moyen et intermédiaire (entre hygrophile et xérophile, avec les combinaisons possibles : mésoxérophile, etc.).
Monophage : Qui se nourrit exclusivement aux dépends d’une seule espèce végétale (antonymes : oligophage et polyphage).
Montigène : habitant des systèmes collinéens ou montagnards.
Multivoltin (entomologie) : Qui possède plusieurs générations annuelles.
Myrmécophile : Dont la larve vit en symbiose associative avec des fourmis.
Nitrophile : Qui affectionne les sols riches en substances azotées (tels les orties des lieux surfumés).
Nymphe : Troisième état du cycle d’un insecte (synonyme : chrysalide).
Oligophage : Se nourrissant d’un nombre très restreint de plantes-hôtes.
Ombrogène : Généré par les pluies.
Opportuniste : tous les êtres vivants le sont puisque tirer le meilleur profit des ressources est la clé de la survie, mais pour certaines espèces, la méthode devient paroxystique. Il s’agit alors d’espèces opportunistes, ou providentialistes, en permanence à l’affût des meilleures conditions possibles pouvant générer une fécondité exceptionnelle. Elles sont aisément identifiables par leur surpopulation lors des années favorables (synonymes de pluvieuses quand il s’agit d’écosystèmes arides ou steppiques), puis leur « disparition » durant les années peu propices. Ce type démographique selon un taux d’accroissement intrinsèque est indiqué par la lettre « r » (stratèges-r). A l’opposé, le type démographique selon une capacité maximale dans l’espace donné, est représenté par la lettre « K » (stratèges-K) qui indique le point d’équilibre entre le potentiel reproductif et la valence environnementale, Ce sont les espèces d’équilibre.
Orophile : habitant de l’étage altimontain.
Oviposition : Acte de ponte. notamment pour la femelle du papillon (l’organe utilisé est l’ovipositeur).
Pacage : Droit d’usage consistant à laisser paître au bois le bétail (synonyme possible : pâturage).
Patrouilleur : dont le mâle effectue d’incessantes patrouilles à la recherche de la femelle. Certains patrouilleurs ne sont pas nécessairement des territorialistes.
Perchis : Etat d’une futaie composée de perches (stade intermédiaire entre gaulis et futaie).
Pessière : Plantation ou forêt d’épicéas.
Phénologie (en entomologie) : Chronologie du cycle biologique et rythme saisonnier des générations.
Photophobie : Crainte de la lumière.
Photosynthèse : Activité physiologique d’une plante verte transformant les éléments minéraux en éléments organiques par l’action de l’énergie lumineuse.
Phototropisme : Réaction de déplacement orienté (tropisme) déterminée par l’action lumineuse.
Phytophage : Insecte qui se nourrit de végétaux
Phytosociologie : Etude des groupements végétaux, basée sur des critères de composition spécifique.
Pionnier : Qualificatif des premiers états d’une succession.
Plastique : Animal ou végétal capable de s’adapter à une vaste gamme de situations, tolérant tous types environnementaux, donc sans exigences prononcées (= élasticité écologique).
Plurivoltin : Qui vole en plusieurs générations annuelles.
Polyphage : Qui se nourrit d’un large spectre d’espèces végétales (antonyme : monophage).
Praticole : Qui fréquente les prés et les prairies.
Protandrie : Désigne chez certains invertébrés comme les lépidoptères, le phénomène d’ émergence plus tardive des femelles en vue d’une fécondation optimum de ces dernières par les mâles déjà éclos en totalité.
Ripicole : Qui hante les rives des cours d’eau ou des ruisseaux.
Ripisylve : Forêt de rives.
Rupicole : Espèce animale ou végétale qui hante les milieux lapilleux (caillouteux) ou rocheux (synonymes : saxicole ou saxatile)
Rudéral : Littéralement « qui croît dans les décombres ». Se dit d’une espèce à ample permissivité anthropique et acceptant les lieux dégradés, comme la proximité des habitations humaines.
Saprophyte : Organisme se nourrissant de matières organiques en décomposition.
Saproxylique : Organisme se nourrissant de bois en décomposition.
Ségétal : Qui croît parmi les céréales et par extension parmi les cultures (exemples : le coquelicot, le bleuet).
Sciaphile : Qui recherche l’ombre (antonyme : héliophile).
Silicicole : Qui affectionne les sols riches en silice.
Sténoèce : Caractérisé par des exigences écologiques étroites et assez strictes (faible valence écologique) et donc contraint à une localisation en des conditions bien particulières (antonyme : euryèce).
Sylvicole : Qui vit dans les formations boisées.
Symbiose : Association durable indispensable et à bénéfices réciproques.
Sympatrique : Qui partage une même région.
Synécologie : la synécologie étudie les communautés d'êtres vivants et le milieu qui les entoure, c'est-à-dire les rapports qui s'établissent entre les diverses espèces végétales et animales et le milieu extérieur. En synécologie, une unité importante est la biocénose. Elle correspond à une communauté d'êtres vivants qui habitent une portion du paysage et sont adaptés aux conditions de ce milieu.
Syntopie : partage d’un même biotope.
Taxon : Unité taxinomique (espèce, sous-espèce, etc.).
Territorialiste (et adepte du hilltopping, perching, ravining) : qui défend son territoire (pour un mâle...) en investissant un repère électif de surveillance sur la partie la plus élevée d’un relief (= adepte du hilltopping) ou d’un objet au sol (pierre, branche, tige) (= espèce percheuse), ou encore en patrouillant inlassablement une portion définie de ravin (= adepte du ravining)
Thermophile : Qui recherche la chaleur.
Transect : Coupe virtuelle selon un axe spatial déterminé, destinée à représenter schématiquement des informations d’ordre écologique (type de milieu, structure de végétation, espèces présentes…).
Trophique : Relatif à la nutrition.
Ubac : Versant d’une vallée exposée à l’ombre (antonyme : adret).
Ubiquiste : Indifférent aux conditions du milieu et donc répandu partout (synonymes possibles : opportuniste ou éclectique).
Univoltin (ou monovoltin, ou monogoneutique) (entomologie) : Qui n’a qu’une génération par an.
Valence écologique : Amplitude de la permissivité d’adaptation aux milieux.
Vernal : Printanier.
Vicariant : Se dit d’une espèce morphologiquement proche et correspondant à une autre d’une contrée différente. Il s’agit pour la plupart d’espèces géminées et taxinomiquement discutables.
Volateur : Animal ailé (oiseau, papillon) présentant une remarquable aptitude au vol.
Xérique : Se dit d’un milieu sec et plus ou moins aride.
Xérophile : Organisme adapté à la vie dans les lieux secs ou arides.
Xérophyte : Se dit d’une plante adaptée à l’aridité.
Xérothermique : Sec et chaud.
Xérothermophile : Qui affectionne ou recherche les lieux secs et chauds.
Xylophage : Organisme se nourrissant de bois.
Désignation de l’appartenance faunistique de l’espèce
(en partie selon DE LATTIN, 1967) :
Cosmopolite : omniprésent (le plus souvent migrateur) ;
Holarctique : à répartition eurasiatique et néarctique (nord-américaine) ;
Eurasiatique : à répartition paléarctique quasiment globale (européenne et asiatique) ;
Eurosibérien : à répartition comprenant le Sud de la Sibérie ;
Holoméditerranéen : présent sur le pourtour de la Méditerranée ;
Pontico-méditerranéen : habitant de la Méditerranée jusqu’au Nord de l’Asie mineure (Alpes pontiques) ;
Atlantico-méditerranéen : habitant le Sud-Ouest de l’Europe (incluant éventuellement l’Afrique du Nord) ;
Ibéro-maghrébin : à répartition ibérique et nord-africaine ;
Endémique nord-africain : restreint à l’Afrique du Nord ;
Endémique marocain : indigène du Maroc ;
Afro-érémien : confiné aux secteurs présahariens du Nord de l’Afrique ;
Saharo-arabique : réparti sur les secteurs désertiques d’Afrique et d’Arabie ;
Subtropical : provenant des régions chaudes situées entre l’Equateur et les zones désertiques (souvent migrateur) ;
Tropical : à répartition équatoriale (souvent migrateur) ;
« Tu crois pouvoir écraser cette chenille ?
Bien, c’est fait : ce n’était pas difficile.
Bien, maintenant, refais la chenille… »
LANZA DEL VASTO
« Chaque espèce est une merveille: une manifestation de la prodigieuse inventivité et créativité de la vie qui s'est développée pendant des centaines de millions d'années et dont nous sommes issus. Les spectacles de l'épanouissement de la végétation au printemps, le retour des oiseaux migrateurs, sont des éléments qui manquent de plus en plus cruellement à l'humanité contemporaine largement confinée dans le béton urbain. Pouvoir jouir de la beauté de la nature est essentiel à notre équilibre psychique ».
HUBERT REEVES
Dernière mise à jour : 25 janvier 2008